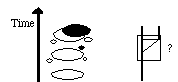
MACROEVOLUTION: TEMPO AND MODE. II
Niles Eldredge and Steven Gould stirred up the mud of Tempo and Mode
in Evolution with their paper in 1972 on so-called "punctuated equilibrium".
The traditional view of evolution was one of phyletic gradualism.
This encompassed slow, gradual change in phenotype and speciation by gradual
change from one species into another. The alternative - punctuated equilibrium
was put forward as a means of accounting for the ever present "gaps"
in the fossil record (see figs. 20.4-20.5, pp. 561-562). Eldredge and Gould
argued that the gaps were not artifacts of incomplete representation,
but that there were essentially no intermediate forms. The general notion
is that long periods of stasis or morphological equilibria
are punctuated by periods of rapid morphological change.
This issue was a bit of a blow to the traditional "Darwinian"
approach to evolution which largely focused on slow gradual change. This
affiliation with "non-Darwinian" evolution is misguided and mislabeled
because the original and updated versions of punctuated equilibrium invoked
speciation in small isolated populations which fits squarely with
Mayr's peripatric model of speciation. Moreover, Darwin described
in the Origin of Species a pattern that is entirely consistent with stasis;
Darwin did believe that the evolution of complex adaptations
was gradual (the eye was built adaptively from preexisting parts in ancestors
and did not pop into being quickly in evolutionary time).
The stratigraphic phenomena would be observed from 1) morphological
stasis in a large population 2) an unrecorded founder event
to a peripherally isolated population 3) speciation, perhaps through
a "genetic revolution", where a new equilibrium morphology would
be assumed and 4) Range expansion of this new form back into the
range of the original form (see diagram below). These events, entirely
consistent with "Darwinian" or "Modern Synthesis" phenomena,
would be observed as a punctuational pattern (see fig. below). Note that
there are other sequences of events that might give rise to the punctuational
pattern.
Several questions arise:
1) What is rapid? 10,000 - 100,000 years can be an instant
in geological time (especially in the context of some deposition
rates) but is ample time for evolutionary events in populations.
Recall that the shift from the peppered to the dark form of Biston betularia
occured within the span of 100 years by a completely "Darwinian"
mechanism.
2) Is rapid morphological evolution associated with
speciation events? Answer: not always (many species of insects
[lacewings, fruit flies] are "good species" but are very difficult
to tell apart). It can be: there are convincing examples of punctuated
patterns in fossil record: Williamson's mollusks.
3) How do we explain stasis? stabilizing selection, developmental
constraints, absence of selection? Eldredge and Gould claim that stasis
is data, i.e., the absence of change is interesting. If stasis
is due to stabilizing selection, then there is perfectly good evolution
going on: selection against individuals at the tails of the distributions
within populations. If stasis is due to developmental constraints then
there is an interesting "battle" going on between the environment
and the homeostasis of the organism. The issue of punctuated equilibrium
has contributed a lot to the science of paleontology since it has
focused new attention on 1) changes at speciation in the fossil record
(see pp. 567-570), and 2) the notion that stasis is interesting and important
and needs explanation.
The publication of the idea of punctuated equilibria set off a bit of
a challenge among paleontologists to show that their "own" mode
of evolution was the correct one. Thus gradualists came out with
papers showing convincing evidence of gradual evolution (figure 20.7, pg.
565) and the Punc. Eq. types came out with papers showing rapid shifts
in phenotype in the fossil record. The absurd example is a data set by
Gingerich which he interprets as gradual and is reinterpreted by Gould
and Eldredge as punctuational! (see below). Like any polarized debate,
there are two kinds of intermediates where reality lies: 1) some data sets
show one mode, others show the other and 2) documented cases of punctuated
gradualism: periods of stasis punctuated by short periods of gradual
change. What remains to be confirmed is whether different lineages tend
to show one pattern and others the other: the relative frequency
of the two alternative modes in the fossil record will ultimately settle
the debate.
The punctuation debated focused a lot of interest on the notion of hierarchical
phenomena (sensu units of selection). One important hierarchical
issue is Species Selection: differential rates of increase or decrease
in species diversity among different lineages due to differences in rates
of speciation and/or extinction. The basic principles of species selection
are 1) speciation is random with respect to phenotype, 2) most changes
occur at speciation, 3) different extinction and speciation rates are due
to some biological properties of the different taxa.
Some consequences: 1) species selection can introduce evolutionary trends
and 2) differences in morphological or taxonomic rates of evolution among
different lineages can be due to species selection. The important point
is that it is the pattern of speciation that drives such trends,
not the direction of morphological changes.
An excellent example of the dynamics of species selection (or how one
might interpret data from the fossil record in light of differences in
extinction and speciation rates) is provided by Hansen's studies of planktotrophic
vs. non-planktotrophic gastropod (snails). Planktotrophic lineages last
longer in the fossil record (lower extinction rate) See fig. 23.3, page
643. However, the proportion of planktotrophs decreases in the fossil
record (see figure 23.4, page 645 and note typo in figure caption). How
can one account for this apparent paradox? If one invokes a higher speciation
rate among non-planktotrophs, then this might do it; i.e., species selection
might account for the patterns of diversity changes. Read the text for
this section (pp. 641-644).
A general question about species selection: is it a pattern or a process? Following the parsimony of G. C. Williams, can we explain species selection by differential survival of individuals within populations, and if so is species selection just a by-product of individual selection., or do higher level processes operate? (thus the hierarchical issue in species selection). If the latter is true, the big question remains: is macroevolution decoupled from microevolution?? (i.e., are population-level processes insufficient to account for evolution above the species level? If you talk to a population geneticist they would say NO! If you talk to a paleontologist some would say OBVIOUSLY!