
PATTERNS OF DIVERSITY
To appreciate the temporal scale of the phenomena that we will discuss,
we should review the geologic time scale and the major eras (Paleozoic,
Mesosoic, Cenozoic) and periods (Cam Or Sil Dev Car Per Tri Jur
Cre Ter Quat) and epochs (Pal Eo Oli Mio Plio Pleisto Holo). The
point here is that there has been immense amounts of time in earth history
(recall our 15 second moment of silence in class as one frame of reference).
Each of these geologic time periods is defined on sedimentary evidence,
much of which is comprised of fossils characteristic of each time period.
On ridiculously simple terms, there has been a dramatic increase in
species diversity from the origin of life to the present. But the patterns
of species diversity are much more complex when viewed in the fossil record.
There are periods of rapid increase in diversity, periods of rapid loss
of diversity (mass extinction events) and subsequent periods of increased
diversity. These patterns beg a variety of questions about the dynamics
of the history of species diversity and what forces might regulate this
diversity: are there equilibria of diversity?, does diversity increase
without limit?, are the patterns of diversity random patterns?
We will first consider some simple models of diversity. Two general processes that affect standing diversity are origination and extinction. These processes are in turn going to depend on the number of taxa, N present at any one time. Thus the change in the number of taxa per unit time, dN/dt can be described as follows:
The rate of origination = O = # new taxa arising / existing taxon / unit time
The rate of extinction = E = probability of extinction during period dt.
thus, change in diversity = dN/dt = ON - EN = N(O-E)
Diversity will be constant if O = E, diversity will increase
if O > E (speciation rates exceed extinction rates) and decrease
if O<E. Diversity might be maintained at a constant level if an
increase in O was matched with an increase in E: is there an equilibrium
diversity? This could only be achieved if there was some feedback
between N, O and E to keep O and E in approximate balance. Consider a graphical
model where the equilibrium number of species depends on the effect of
N on the values of E and O (see figure).
These models just provide ways of thinking about the history of diversity
on earth. What might such diversity dependent forces/phenomena be?
Do they exist?
What do the data tell us? First, how do we get the data? Simple on one hand: count the number of taxa present in each time interval. Complicating issues: species concept = typological (morphospecies) any "reasonably" different forms will be classified as a different species. If these morphospecies co-occur in time and space then they are probably "good" species and should be counted as separate entities. Many fossil series show temporal gradations of morphology from one form into another. These chronospecies introduce error into estimates of diversity since the end of one name and the beginning of another name look like extinction and origination, or pseudoextinction and pseudoorigination. Moreover, the taxononic level under consideration will alter one's estimate of the patterns of diversity since lower taxa (species, genus) might vary in diversity more than higher taxa (family, order). Also, differences in the identification of taxa will alter estimates of patterns (see figure 20.12, pg.577).
In the literature it is difficult to tell which extinction/speciation
events represent real events and not "chrono" events. We can
avoid this problem by tabulating higher taxa. It is unlikely that
all representatives of a genus will evolve simultaneously into a new genus,
but what is a genus (or a family)? is it a real unit of diversity?
Additional biases in quantifying diversity: thickness of sediments
different sediments deposit at different rates so that the resolution
is quite different among sediments (affecting the rate (dt) component
of the dN/dt calculations). Area of exposed sediments are not always
representative of the temporal and spatial scales one might want to sample/represent
in a fossil assemblage.
Additional artifact of the age of the rocks known as "the pull
of the recent" Young rocks are less likely to be destroyed by
the forces of time than are old rocks. This means that last occurrences
of a fossil are more likely to be better recorded than first
appearances (actual first "appearance" might have been exposed,
eroded, reworked, etc. moving the observed first appearance up in
time). Thus the duration of lineages will be "pulled" toward
the recent (pulled implies an artifact of increased likelihood of a fossil
to be recorded in younger rocks). Similarly, and extant group with
a single fossil occurrence has a temporal range from its fossil date through
to the present, but and extinct group with a single fossil occurrence
has only that single date as a temporal range, even if it lived for a long
time but was not deposited as a fossil.
Biases aside, what does the record tell us? Does diversity increase
with time? The Cambrian Explosion ("sudden" appearance
of many invertebrate phyla in the early Precambrian) shows a clear patterns
of increase in diversity from the known forms present in Pre-Cambrian strata.
Were there some physical triggers of diversity increase such as
geologic activity inducing major changes in the physical environment?.
Or were there some biological triggers that allowed the rapid evolution
of new forms? These might consist of the evolution of a key innovation
(a unique new trait that allows a species/taxon to exploit a new way of
life)? This idea invokes the concept of the adaptive zone,
a new set of ecological niches utilized/exploited by a group of related
species in which this group might radiate (speciate) extensively and increase
in diversity.
The diversity of marine invertebrates in the Phanerozoic
(period of visible life) is the best example. Increase from Cambrian
to Ordovician, then mass extinction; increase to plateau through
the Paleozoic (major extinction events in middle of plateau); continuous
increase from Triassic through Tertiary (see figure below).
Patterns of increase of diversity indicate three evolutionary faunas:
1) Cambrian, 2) Paleozoic and 3) Modern (work of J. Sepkoski). Despite
the major patterns, different taxa have very different profiles: there
are examples of extinction and reradiation, maintenance of
diversity and continuous diversification all spanning the same time
frame (compare fig. below with other taxa: fig. 21.11 - 21.16. pp. 602-607).
This suggests that if there are biological forces/pressures regulating
diversity, they may be acting quite differently in different taxa.
Increases in diversity were measured by Bambach in a slightly different way: he classified organisms into guilds (groups of organisms with similar ways of life) and examined the increase in taxonomic diversity from the perspective of guilds. Conclusion: increase in diversity through the Phanerozoic achieved more by the increase in number of guilds rather than increasing number of species within each guild. This suggests that "ecospace" becomes more tightly packed through the Phanerozoic. Ecospace is represented by three axes and different species would "fill" different regions of the "cube" of ecospace.
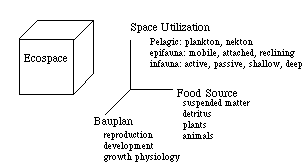
All these observations again beg the question: is diversity regulated,
or put another way, can we treat the patterns of diversity in the fossil
record as we might treat an ecological system? There are inferences
that support both answers to this question. Computer models where
the rates of extinction and origination (speciation) fluctuated randomly.
One constraint was that the mean probability of a taxon going
extinct equaled the mean probability of it speciating.
Thus, diversity would fluctuate but hover about some "equilibrium"
value. Results produced patterns of diversity within clades very similar
to those observed in the fossil record (see below).
Data supporting the notion of regulation is the observation that rates
of extinction and rates of origination are correlated.
This suggests that when speciation increases diversity, extinction rates
increase and bring diversity back down; similarly if extinction rates increased,
speciation rates would increase and bring diversity back up (beware of
using a teleological argument; these changes are effects
of changes in diversity presumably occurring due to ecological pressures
at the time, not for the purpose of restoring diversity). Observation
of correlation of extinction and speciation rates suggest diversity
dependent effects or feedback in an ecological sense.
Can also model patterns of increase in diversity as one would model exponential growth in ecology Nt = N0ert where r = birth rate - death rate (or in our case, origination rate - extinction rate). N0 = 1 species (beginning of group) and t = time since first fossil
The data from Cambrian fit this relationship fairly well. Diversity
during the Paleozoic fit a Logistic growth curve quite well (logistic
curve has a feedback where number if individuals [taxa] levels off with
increasing density; sigmoid curves in figure below)
One can extend this analogy even further and treat Sepkoski's three
evolutionary faunas as one might treat competing species. The patterns
fit models of species competition. But: the observation is a pattern
in the fossil record and we do not know whether the process of competition
existed between the two major faunas. This pattern can be seen on a lower
taxonomic level also: rodents increase in diversity as the multituberculates
(rodent-like mammals) decrease to extinction. Shortly after the brachiopods
(clam-like marine invertebrates) bite the dust, the bivalves (e.g.,
clams) diversify extensively. The patterns look like what one might expect
from competitive exclusion, and is referred to as ecological
replacement since one group with a similar set of key innovations replaces
another group (see figure below).
Re analysis of the brachiopod/bivalve replacement has suggested that there has been no interaction between the "competing" forms and that they are best though of as "ships that pass in the night". This does not mean that all ecological replacements in the fossil record do not involve competition, just that it is hard to say. A nice way to compare the possibility of interaction, or the lack of it, is illustrated in figure 21.12, pg. 603.