SEXUAL SELECTION
So far all of our discussions of selection have been without much regard
for the sex of the individual under selection (evolution of the sex ratio
to 1:1 can occur through selection for alleles in either
males or females that favor the production of the rare sex). But
just as with natural selection individuals may differ in their ability
to reproduce, in sexual selection there can be differential reproductive
success among individuals of the same sex (and species). In order to mate,
males need to gain access to females and vice versa, and not all individuals
will be equally successful at this task. If there are genetically based
differences in the ability of one sex to insure successful mating with
the other sex, sexual selection will occur.
The patterns and processes of sexual selection are best understood in
the context of parental investment. Sexual selection occurs because
there is a correlation between the gender of an individual and its parental
investment in each offspring. Parental investment is the investment
of resources that increases in the probability offspring will survive
(a benefit) while decreasing the parent's ability to produce more
offspring (a cost). By investing in the production of current offspring
a parent will reduce the likelihood that it will be able to invest
in future offspring. Costs can be the energetic demands of parenting, increased
predation risk, etc.
Parental investment should be proportional to:
(Benefit to current offspring survival) / (Cost to future offspring survival)
One important component of parental investment is the investment
in gametes. This serves as one means of distinguishing the sexes. Females
are the sex with a large parental investment per gamete. Example
are eggs which have the nutrients to promote the development once fertilization
occurs. Males have a smaller parental investment per gamete;
generally carries only genetic information.
Some birds: each egg = 15-20% of a females body weight. In males,
millions of sperm produced each ejaculation; total output per reproductive
season < 5% of body weight. (The difference is less pronounced,
but still significant, in mammals). This difference in investment in gametes
has important consequences for how these resources are invested
and hence how selection might act differently in the two sexes. In general,
females should commit their eggs for reproduction prudently; males
need not be so cautious in their commitment of sperm. Put in evolutionary
terms, the reduction in fitness of a female that squanders an egg will
be greater than the reduction in fitness of a male that squanders a sperm
(give or take a million).
Another important component of parental investment is parental care,
care of the zygote after fertilization. The amount of parental care is
also frequently quite different in males and females. In species with internal
fertilization the female is "stuck" with the egg after fertilization.
In many species the female also cares for the young after they hatch. Males
are not anatomically tied to the egg and hence are sometimes "freed"
from this additional parental care.
Moreover, in species where multiple matings occur, a male cannot
be certain that a females offspring were fathered by him. Benefit
of caring for unrelated offspring is low, a male can increase
fitness by avoiding parental care.
General result is that sex with higher parental investment = limiting
resource. Since this sex is usually females, this will lead to male-male
competition for access to mates (like resource competition in ecology),
and female choice where females choose among males so that they
may prudently commit their higher parental investment.
Put another way, males will increase fitness by increasing the
number of fertilizations they can perform, leads to Intrasexual
selection (selection among individuals of the same sex). Females will
increase fitness by being choosy, leads to Intersexual
selection where the high-investment sex chooses among the low investment
sex.
As a consequence, these forms of sexual selection can lead to the evolution
of traits that better enable each sex to perform its "fitness-increasing"
behavior. In males: Horns, large body size, sperm competition, mating
plugs. In females: choosy behavior . The existence of choice
in females can lead to traits in males that tend to give individuals an
advantage in attracting mates: Plumes, coloration. These present
a problem, however. Have these phenotypic characters evolved so that males
can out compete other males to gain access to females, OR, have they
evolved so that a male might win and the be in a position to be chosen
by females? The relative contributions of intra and intersexual selection
in the evolution of some traits can be difficult to distinguish.
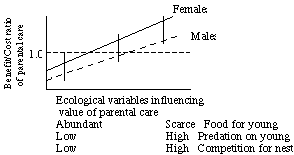
Ecological contexts in which parental care might be given can influence the amount of parental care invested in offspring.
For species in the lower left of the graph it doesn't pay either sex
to invest in parental care; species in the middle should exhibit
sexual dimorphism (male showy, female less so or cryptic to reduce
predation risk); species at the upper right should both exhibit
parental care, monomorphic for plumage characteristics in birds
Evidence that the certainty an individual has regarding his/her paternity/maternity influences the amount of parental care is available in fishes and amphibians. The table presents the numbr of genera in each group (fish, amphibians) that fall into the categories.
| Sex: | Male Parental Care | Female Parental Care | ||
| Fertilization: | Internal | External | Internal | External |
| Group | ||||
| Fishes | 0 | 48 | 15 | 21 |
| Amphibians | 2 | 13 | 11 | 7 |
Internal fertilization: male cannot be sure that his sperm
will fertilize the eggs in the female; another male could come along and
mate the female, displacing his sperm parental care does not pay.
With external fertilization a male can be quite certain that some
of his sperm will fertilize eggs so parental care does pay
Examples of sex role reversal: male has higher parental care,
is the high investment sex and should choose among females; females
are the showy sex, males are the cryptic sex (another bird example
= phalaropes, female is showy). Difficulties of determining what is
being chosen: good genes?, good gifts? (wasp example) what is it about
a male that will bring high fitness to one's offspring?
Zahavi handicap model (a peacock's tail is a handicap because
it may reduce fitness by natural selection): females choose males
with handicaps because males with handicaps must have "good genes"
in order to be exant while carrying the fitness-reducing handicap. Such
a system might evolve if the female's advantage by mating with the "good
genes" of this male outweighed the cost of her offspring having to
carry this handicap around.
Kirkpatric model: flashy trait in male and the preference for it in female will become associated (in technical terms, the alleles for the male's flashy trait and the alleles for females choosing this trait come into linkage disequilibrium). Runaway sexual selection results which can be non adaptive; sexual and natural selection can oppose one another.