
LEVELS OF SELECTION
For natural selection to proceed there must be heritable variation
in phenotypes and the variation in phenotype must be associated with differential
survival and/or reproduction, i.e., there must be differential fitness.
By inference then, any entities exhibiting heritable variation in
rates of reproduction can evolve. We need not restrict our thinking
to "individuals" in "populations" in the traditional
senses of these words.
Nature is organized in a hierarchical fashion. In terms
of entities that can be heritable we can consider genes, chromosomes,
genomes, individuals, groups, demes, populations, species, etc. Each
of these entities meets the requirements of units that can be acted upon
by selection. At which level(s) does selection act? Answer: all of them.
What then is the important unit of selection? Answer: it depends.
First, some historical context. Serious consideration of a unit of selection
other than the individual was advanced by V. C. Wynne-Edwards (1962, Animal
Dispersion in Relation to Social Behavior). Populations have their
own rates of origination and extinction and selection could thus operate
at the level of the group. Idea based on observation that many species
tend to curb their reproductive rate/output when population densities are
high. This behavior would favor groups that exhibited the behavior and
select against those that did not; i.e., there would be group selection.
G. C. Williams responded to this idea with Adaptation and Natural
Selection (1966) arguing that this behavior would be less fit than
a cheating behavior where individuals did not reduce their reproductive
output at times of high density/low food availability. In general selection
at the level of the individual would be much stronger than selection at
the level of groups. In keeping with Williams' claim that one should always
seek the simplest explanation for selective/adaptive explanations,
individual selection is usually sufficient to account for patterns.
Group selectionist thinking leads to statements such as "good for
the species" when it is entirely likely that it may be good for the
individual as well: reduced reproductive effort in times of low food may
increase an individual's reproductive output at a later date.
Examples of selection at different hierarchical levels: Genic selection
is selection at the gene level; best example is meiotic drive (segregation
distortion) where one gametic type (often one chromosomal type) is
transmitted into the gamete pool (or next generation) in excess (or deficiency).
The T locus in mice: affects tail length but also viability. TT homozygotes
have normal long tails; Tt heterozygotes have short tails and transmit
~ 90% of the t allele to their sperm; tt homozygotes are sterile. Meiotic
drive will increase frequency of t allele to point where that become frequent
enough to occur as tt homozygotes with appreciable frequency, whereupon
selection works against t alleles. Opposite Selection at two levels
selection for at the level of the gene; against at the level
of the genotype (organism).
In this system the balance of opposing selection coefficients at different
levels should give an equilibrium allele frequency of 0.7 for f(t) allele
(using data not provided here). In nature f(t) = 0.36. Discrepancy due
to small local groups and drift. Some local breeding groups (2 -
4 individuals) fixed for the t allele and since tt is sterile, these demes
go extinct reducing the f(t). Thus we have selection at three levels:
genic, individual and intergroup all contributing to the maintenance of
the t/T polymorphism.
Would we expect to detect meiotic drive systems in natural populations?
If a new mutation arose that introduced a bias in the transmission of the
chromosome on which it was located, then it would sweep to fixation and
the locus would be homozygous for the "drive" allele. Meiotic
drive can only be detected in heterozygous state, so the
drive system would disappear when the drive allele went to fixation. There
will be a window of time where the allele is increasing in frequency, but
this could be short-lived. If the drive allele reduced viability
in the homozygous state (as the T locus example), then variation can be
maintained and the drive system would persist longer, making it more likely
to be detected.
Another case where genic selection may act: sex ratios. Why should the
sex ratio be 1:1 in most diploid species? Assume a sex ratio of 40% males
and 60% females. Males in this case are in limited supply. Any gene leading
to the production of more males (a allele results in more males the A allele
at a sex determination locus) will be favored until the frequency of males
is >50%. Sex ratios tend to stabilize at 50:50 (R. A. Fisher, 1930).
Kin selection and altruistic behavior many species of animals
that live in groups give warning calls which alert other individuals
about predators, etc. How could this behavior evolve when making the call
alerts the predator to the callers location and increases the possibility
of the caller becoming prey. Mammals that nurse their young: major energy
investment for the mother may be thought to reduce her fitness, but make
obvious sense sine the individuals that benefit are close relatives (offspring).
Put in fitness terms: how could a trait evolve that lowers individual
fitness?
Key point is the term individual. If the ones that benefit from
the behavior are related, the loss in individual fitness may be
regained in inclusive fitness, i.e., individual fitness plus fitnesses
of relatives. W. D. Hamilton argued that an altruistic trait could
evolve if cost to altruist/benefit to recipient < genetic relatedness
(C/B < r) where r is an estimate of the probability of the donor
and recipient having allele identical by descent. For example parent -
offspring have r=0.5; siblings have r=0.5; grandparent-grandchild have
r=0.25.
Idea of inclusive fitness implies that one's fitness is determined
by one's own life time reproductive output and the reproductive
output of relatives, scaled by their degree of relatedness (r). In the
warning call example, if calling out to warn about the arrival of a hawk
killed you but saved three reproductively active siblings, it would have
been worth it. If it only saved two siblings, it probability wasn't worth
it. Obviously one cannot tabulate the payoff of event x, y, or z. The point
is that the notion of inclusive fitness provides a fitness context where
altruistic behavior could evolve even when it appears to decrease individual
fitness. Two modes of selection: individual selection opposes
altruism; selection among kin groups favors altruism.
Classic examples: helpers at the nest in birds. Young offspring
remain at the nest to help their parents produce more siblings in subsequent
years/seasons. Helpers may contribute more to their own fitness by aiding
in the production of siblings than by trying to reproduce themselves and
failing due to lack of experience or availability of nest sites. Sterile
workers in hymenoptera (ants, bees, wasps): males are haploid (develop
from unfertilized egg) so sisters have r=0.75 because male contributes
the same allele (relatedness between sibs for paternal genes = 1.0;
relatedness between sibs for maternal genes = 0.5; among diploid
female workers this averages out to r=0.75). A female worker does more
to propagate her own genes by staying in the nest and aiding in the production
of sisters (r=0.75) than by going off and producing her own daughters (r=0.5).
Used as an explanation for the evolution of sociality (e.g., colonies)
in hymenoptera.
Group selection = variation in the rate of increase or extinction
among groups as a function of their genetic composition. Again consider
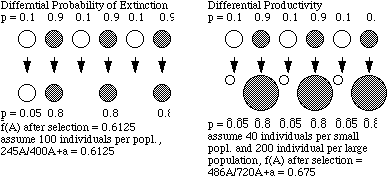
how an altruistic trait could increase in frequency. Differential rates
of extinction: allele A confers altruistic behavior; at a selective
disadvantage to allele a within the group. Should lead to the reduction
of allele A. But groups with high frequency of A may be less likely to
go extinct (due to better exploitation of resources). Over all groups with
high frequency of A persist and f(A) increases.
Differential productivity: similar to model above but altruistic
trait affects reproductive output of group. Selection against A
allele within groups (selfish types have higher short term fitness) but
groups with high frequency of A exploit resources more prudently and actually
produce more offspring over the long term: f(A) increases. Model this as
follows: assume a haploid trait with A=altruist, a=selfish; p=f(A).
In each population p decreases within a population through time
due to selfish individuals out competing altruistic individuals. But in
all populations as a whole the altruist gene increases over time.
Below, the average f(A) across all populations is 0.5 at the start:
Clearly, if this system were to continue for many generations, the frequency
of the altruist gene would decrease within each population. But under conditions
where the selection favoring selfish genes was weak and the group selection
increasing the probability of staying extant (or the growth rate of the
population) was strong, and altruist allele might be preserved. Because
the conditions are so restrictive, group selection is presumed to be a
rare phenomenon.
Group selection often involves plausible models but require that interdeme
(group) selection be strong. Would have to be very strong to overcome
selection among individuals within populations. Other complicating factors:
turnover rate of individuals is faster than of populations/groups;
fixation of less fit allele is unstable to invasion by new mutant
allele or "selfish" allele introduced by gene flow. New research
on multilevel selection suggests that there should not be the necessary
association between altruism and "sacrifice" or genetic "suicide".
Cooperation among individuals can actually result in higher group fitness
without the assumed loss of individual fitness (see a meeting review in
Science (9 August 1996) vol 273:739-740). D. S. Wilson makes the
analogy between the optimal clutch size argument of D. Lack and the optimal
group of Wynne-Edwards. With too many eggs in a clutch an individual may
die trying to support them all, so some intermediate clutch size is "optimal"
(see Life History Lecture). Optimal groups may evolve intermediate density
by the same trade-off mechanism.
A further problem for group selection: with localize population structure,
there can be considerable inbreeding which increases relatedness
(r). Thus inter "group" selection that gives the appearance of
evolution of altruistic traits may be mediated by kin selection
due to the high relatedness among individuals.
Later we will consider species selection. Some lineages have
more species than others, but are these lineages more fit? Is this simply
a pattern (more species) or is it really a different process?
Is it simple like bacteria in chemostats: a higher birth/death ratio; some
lineages seem to speciate faster than their members go extinct? Is this
mediated at the level of the species, or can we explain it (as G. C. Williams
might like) at the level of individuals within populations?
Richard Dawkins likes to couch this discussion in terms of replicators and vehicles. Replicators are any entities of which copies are made; selection will favor replicators with the highest replication rate. Vehicles are survival machines: organisms are vehicles for replicators and selection will favor vehicles that are better at propagating the replicators that reside within them. There is a hierarchy of both replicators and vehicles. The key issues are that 1) the "unit" of selection is one that is potentially immortal: organisms die, but their genes could be passed on indefinitely. The heritability of a gene is greater than that of a chromosome is > that of a cell > organism > and so on. But , because of linkage we should not think of individual genes as the units; it is the stretch of chromosome upon which selection can select, given certain rates of recombination. Issue 2) is that selection acts on phenotypes that are the product of the replicators, not on the replicators themselves, but the vehicles have lower heritability and immortality than replicators. What then is the unit of selection?? All of them, just of different strengths and effects at different levels.