FITNESS AND ADAPTATION. I
General observation: most organisms show "good fit" to their
environment. Desert plants (desiccation resistant), marine organisms
(fusiform body), cryptic insects All generally can be placed in
the context of the environment presenting a problem and the organism,
through evolutionary changes, providing a solution to the problem
posed by the environment. The apparent "good fit" between organism
and environment suggests that this state increases the survivorship and/or
reproduction of the organism.
This may lead us to identify certain problems posed by the environment
and ask how well does the organism solve the problem Leads to a
design/engineering approach to defining survival ability, design
fitness or engineering fitness. We do this by comparing form and function;
often there is an "appealing" match or sense of "fit"-ness.
Is this what we mean by fitness
We then ask if the design/solution is the optimal solution suggesting
an absolute engineering fitness scale where the trait/attribute
is assessed with respect to an ideal solution to the problem. Alternatively,
comparing different available solutions may be all that matters:
a relative engineering fitness
This can be difficult because we could almost always "design"
a system or part of an organism with a slight improvement over what it
has achieved by evolution. Does the presence of a design that "appears"
to have room for improvement mean that it is not "fit" Need to
be aware of the design constraints of organisms before making useful/contributory
statement about level of engineering fitness.
Darwinian fitness is assessed not by looking at the "fit"
between form and function, but by attempting to determine the contribution
to future generations. What about grandchildless mutants in
Drosophila: have normal number of offspring, but these offspring are sterile.
How many generations ahead do we need to look? Since we can never
really look ahead can we really identify fit individuals/genotypes?
Another old problem: is Darwinian fitness a tautology? (a statement
that is true by virtue of its logical form; needless repetition of an idea),
e.g. my father is a man; Brown students are smart; the water is wet. Natural
selection was characterized by Huxley as "the survival of the fittest".
The allele with high fitness will increase; allele A has high fitness;
thus allele A will increase in frequency. The premise provides for no alternative
outcome, therefore it is a useless theory. Survival of the fittest; who
are the fittest? those that survive. Thus it reduces to survival of
the survivors. Characterized as such this tell us nothing and cannot
be predictive?.
How do we get out of this? Assign the predictive aspect to the
term fitness. Distinguish between realized vs. expected fitness.
A population passes through a generation, look at frequencies of genotype
before and after transition, define fitnesses by changes in frequencies.
This approach to fitness can lead us into the tautology trap: fitness defined
by those that survive better (post hoc), but the change may
not have been due to natural selection (e.g. drift). With true
Darwinian fitnesses we would be able to predict which genotype
would survive better (fitness values assigned to genotype "before"
generation transition). The "fitter" genotype would have an inherent
tendency to out reproduce competing genotypes.
G. C. Williams in Adaptation and Natural Selection, 1966, Princeton
U. Press, states that the problem "is not survival as such, but design
for survival" (pg. 159). Thus overall "fitness" is a combination
of Darwinian and engineering fitnesses.
How do we measure fitness? Bacterial chemostats, one genotype increases
at the expense of the other. Fine if we believe that environments are ~constant
over time (maybe they are in the intestine) and that alleles with different
fitnesses can be considered outside the context of the entire genotype.
Probably unrealistic for many haploids (and most diploids).
In diploids a bit more difficult: is it the phenotype, genotype or
allele that determines fitness. Well it is all: selection acts on
phenotype which is produced by a genotype in a given environment
leading to the change in allele frequencies which in the next generation
gives different genotype frequencies. Problem: neither genotype or phenotype
are inherited intact (phenotypic "inheritance" depends on heritability
of trait VG/VP )
One approach is to determine fitness of alleles e.g. "average excess" of allele:
p' = (p2 wAA + pqwAa) / (p2 wAA
+ 2pqwAa + q2waa) = [p (pwAA
+ qwAa)] / (p2 wAA + 2pqwAa +
q2waa)
where pwAA+ qwAa = wA = fitness of A allele. Thus p' = p
Think about it: this is the frequency with which the A allele pairs with itself (the A allele) resulting in fitness wAA plus the frequency with which the A allele pairs with the a allele resulting in fitness wAa relative to (i.e., divided by) the average fitness in the population (wbar). This is a very nice formula since the ratio wA/wbar determines whether p will increase or decrease in the next generation.
Fitness (selective) values of genotypes are a similar thing since they
average the interaction effects of the AA genotype over all other genotypes
at all other loci in the genome for a given environment: average fitness
of AA when coupled with the B locus: (wAA = p2wAABB
+ 2pqwAABb + q2wAAbb ) assuming p=f(B)
and q=f(b).
Note: selection will continue until: wA = wbar (i.e., the
ratio is = 1.0 and p' = p(1), no change). Thus average fitness in
the population will be maximal but this does not mean the fitness of
the population is maximal.
Need to distinguish between individual fitness and population fitness.
wbar is a description of the average fitness of all individuals in the
population. Does this determine the fitness of the population? If
we think of the population as the unit of "survival" a population
with high fitness is one that has a high tendency to out reproduce other
populations. This is largely a demographic question since drought,
predators, parasites, etc. could alter a populations' probability of survival
irrespective of the number of individuals it is producing each generation.
A highly fit population is one that has a high reproductive output or
is unlikely to go extinct. These qualities can be unrelated to the average
fitness in the population (although they can be related).
Consider wasps living in nest holes. Population is limited
by a fixed number of nest holes. Now a mutation occurs that doubles
the egg output of individuals carrying the mutation. Those individuals
with mutation are more fit, but the population will not increase due to
limitation of number of nest holes.
Consider prey switching in birds: mutation increases the fecundity of
a moth. More caterpillars present, birds switch their search image and
prey more heavily on the abundant caterpillars: reduces population size
clearly the population is less fit.
These examples have assumed that population size is some measure
of average fitness May not be the case depending on the mode of selection:
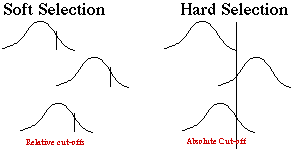
Soft versus hard selection. Soft selection is like grading on a
curve: there are always a certain number of As (or F's). Regardless of
the absolute fitness of individuals or the population the "better"
ones will survive to fill those spots. Hard selection assumes some threshold
level of "fitness" that could be set by the environment. If all
individuals are below this level, population crashes; if all are above,
population grows. See figures 7.4, 7.5, pgs. 166 - 167) .